Los mecanismos enzimáticos individuales se han deducido mediante una diversidad de métodos, que incluyen experimentos cinéticos, estudios estructurales de proteínas y análisis de reacciones con modelos no enzimáticos. Los resultados de esos estudios indican que la extraordinaria capacidad catalítica de las enzimas se debe a propiedades físicas y químicas sencillas, especialmente el enlazamiento y la colocación adecuada de los sustratos en los sitios activos de las enzimas.
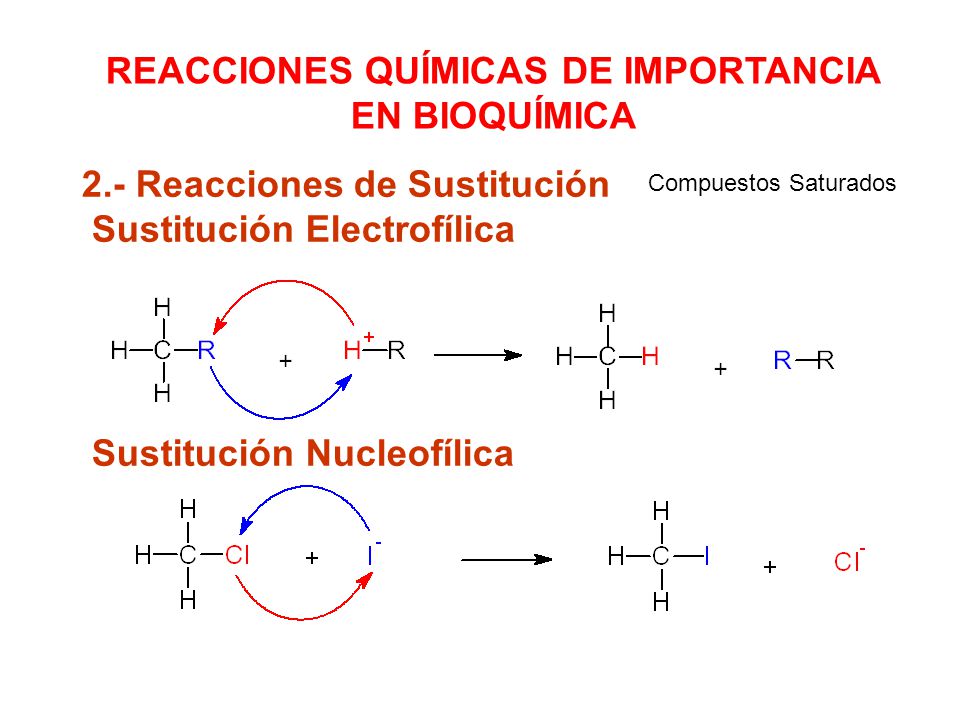 Sustituciones nucleofílicas
Sustituciones nucleofílicas
Muchas reacciones químicas tienen compuestos intermedios iónicos. Hay dos tipos de
intermedios iónicos: unas especies son ricas en electrones o nucleofílicas, y otras especies
son pobres en electrones, o electrofílicas.
Un nucleófilo tiene una carga
negativa o un par de electrones no compartido. Suele imaginarse que el nucleófilo ataca
al electrófilo, y al mecanismo se le llama ataque nucleofílico, o sustitución nucleofílica.
Reacciones de ruptura
También se encuentran reacciones de escisión o ruptura. Los enlaces covalentes se pueden
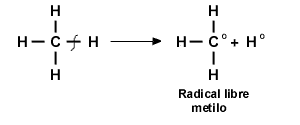 romper de dos maneras: dos electrones permanecen con un átomo, o un electrón puede permanecer con cada átomo enlazado. En la mayor parte de las reacciones, ambos electrones están en un átomo, por lo que se forman un intermedio iónico y un grupo saliente. Por ejemplo, la ruptura de un enlace C—H casi siempre produce dos iones. Si el átomo de carbono retiene ambos electrones, el compuesto que lo contiene se transforma en un carbanión.
romper de dos maneras: dos electrones permanecen con un átomo, o un electrón puede permanecer con cada átomo enlazado. En la mayor parte de las reacciones, ambos electrones están en un átomo, por lo que se forman un intermedio iónico y un grupo saliente. Por ejemplo, la ruptura de un enlace C—H casi siempre produce dos iones. Si el átomo de carbono retiene ambos electrones, el compuesto que lo contiene se transforma en un carbanión.
Reacciones de oxido-reducción
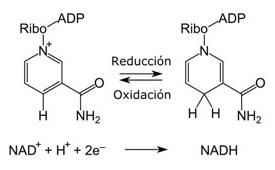 Las reacciones de oxido-reducción son fundamentales en el suministro de energía biológica. En una reacción de oxido-reducción (o reacción redox), los electrones de una especie se transfieren a otra. Aquí la terminología puede ser bastante confusa, por lo que es importante dominar el significado de las palabras oxidación y reducción, pues aparecerán en forma repetitiva en el resto del libro. Oxidación es la pérdida de electrones: una sustancia que es oxidada tendrá menos electrones cuando se termine la reacción. La reducción es la ganancia de electrones: una sustancia que gana electrones en una reacción es reducida. Las reacciones de oxidación y reducción siempre suceden en forma simultánea. Un sustrato es oxidado y el otro es reducido. Un agente oxidante es una sustancia que produce una oxidación: toma electrones del sustrato que es oxidado. Así, los agentes oxidantes ganan electrones (es decir, se reducen). Un agente reductor es una sustancia que dona o cede electrones (y en el proceso se oxida).
Las reacciones de oxido-reducción son fundamentales en el suministro de energía biológica. En una reacción de oxido-reducción (o reacción redox), los electrones de una especie se transfieren a otra. Aquí la terminología puede ser bastante confusa, por lo que es importante dominar el significado de las palabras oxidación y reducción, pues aparecerán en forma repetitiva en el resto del libro. Oxidación es la pérdida de electrones: una sustancia que es oxidada tendrá menos electrones cuando se termine la reacción. La reducción es la ganancia de electrones: una sustancia que gana electrones en una reacción es reducida. Las reacciones de oxidación y reducción siempre suceden en forma simultánea. Un sustrato es oxidado y el otro es reducido. Un agente oxidante es una sustancia que produce una oxidación: toma electrones del sustrato que es oxidado. Así, los agentes oxidantes ganan electrones (es decir, se reducen). Un agente reductor es una sustancia que dona o cede electrones (y en el proceso se oxida).
Modos químicos de la catálisis enzimática
La formación de un complejo ES coloca a los reactivos en la cercanía de residuos de aminoácidos del sitio activo de la enzima. Las cadenas laterales ionizables participan en dos clases de catálisis química: catálisis ácido-base y catálisis covalente. Son los dos modos químicos principales de catálisis.
Residuos polares de aminoácidos en sitios activos
La cavidad del sitio activo en una enzima en general está cubierta con residuos hidrofóbicos de aminoácidos. Sin embargo, también en el sitio activo hay unos pocos residuos polares, ionizables (con algunas moléculas de agua). Los residuos polares de aminoácidos (o a veces coenzimas) tienen cambios químicos durante la catálisis enzimática. Esos residuos forman gran parte del centro catalítico de la enzima.
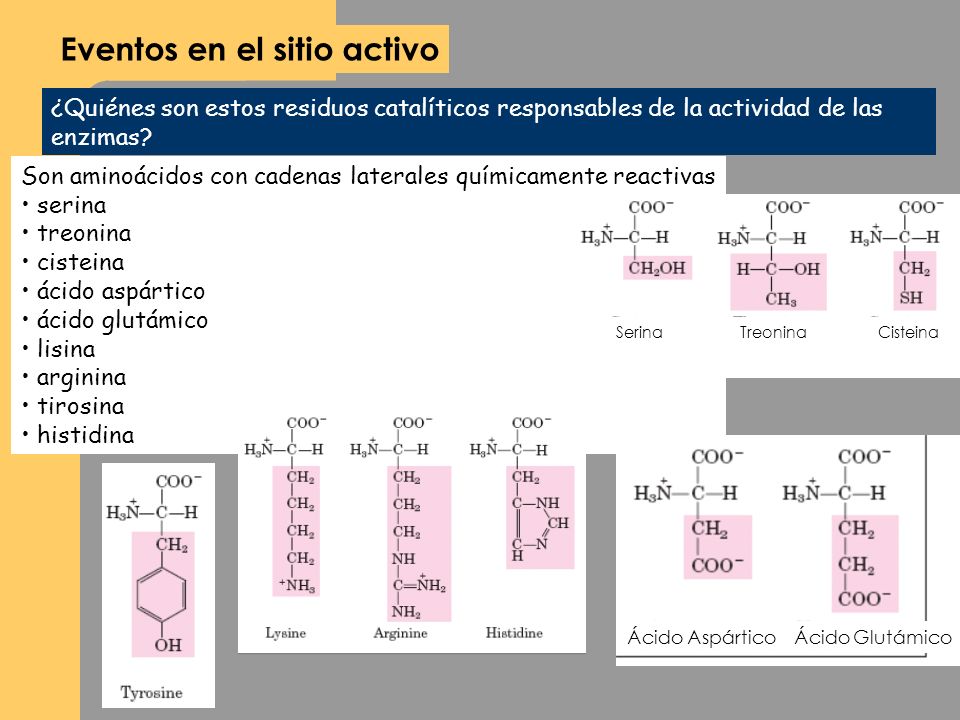 Las enzimas suelen tener de dos a seis residuos catalíticos esenciales. Los diez residuos catalíticos más importantes se muestran en la imagen Los residuos con
Las enzimas suelen tener de dos a seis residuos catalíticos esenciales. Los diez residuos catalíticos más importantes se muestran en la imagen Los residuos con
carga, His, Asp, Arg, Glu y Lys forman casi las dos terceras partes de todos los residuos catalíticos. Eso tiene más sentido, porque es más probable que las cadenas laterales con carga funcionen como ácidos, bases y nucleófilos. También es más probable que tengan un papel en el enlazamiento con el sustrato, o con estados de transición. La histidina es el residuo catalítico número uno. Es seis veces más probable que intervenga en la catálisis que lo que parecería indicar su abundancia en las proteínas.
Catálisis ácido-base
 En la catálisis ácido-base, la aceleración de una reacción se debe a la transferencia catalítica de un protón. Esta catálisis ácido-base es la forma más común de catálisis en química orgánica, y también es común en las reacciones enzimáticas. Las enzimas que la hacen se basan en cadenas laterales de aminoácido que pueden donar y aceptar protones en las condiciones de pH neutro de las células. Este tipo de catálisis ácido-base, donde intervienen agentes de transferencia de protones, se llama catálisis ácido-base general. (La catálisis por H u OH se llama catálisis ácida específica, o catálisis básica específica). De hecho, los sitios activos de estas enzimas son el equivalente biológico de una
En la catálisis ácido-base, la aceleración de una reacción se debe a la transferencia catalítica de un protón. Esta catálisis ácido-base es la forma más común de catálisis en química orgánica, y también es común en las reacciones enzimáticas. Las enzimas que la hacen se basan en cadenas laterales de aminoácido que pueden donar y aceptar protones en las condiciones de pH neutro de las células. Este tipo de catálisis ácido-base, donde intervienen agentes de transferencia de protones, se llama catálisis ácido-base general. (La catálisis por H u OH se llama catálisis ácida específica, o catálisis básica específica). De hecho, los sitios activos de estas enzimas son el equivalente biológico de una
solución de ácido o base.
Catálisis covalente
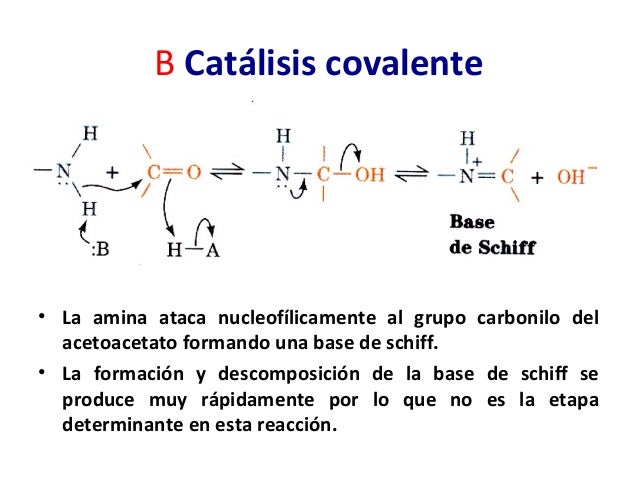 En la catálisis covalente se une un sustrato en forma covalente a la enzima y se forma un compuesto intermedio reactivo. La cadena lateral que reacciona de la enzima puede ser un nucleófilo o un electrófilo. La catálisis nucleofílica es más común. En el segundo paso de la reacción se transfiere una porción del sustrato del compuestointermedio a un segundo sustrato. Este es un mecanismo común en bioquímica para acoplar dos reacciones diferentes. Recuérdese que la capacidad de acoplar reacciones es una de las propiedades importantes de las enzimas
En la catálisis covalente se une un sustrato en forma covalente a la enzima y se forma un compuesto intermedio reactivo. La cadena lateral que reacciona de la enzima puede ser un nucleófilo o un electrófilo. La catálisis nucleofílica es más común. En el segundo paso de la reacción se transfiere una porción del sustrato del compuestointermedio a un segundo sustrato. Este es un mecanismo común en bioquímica para acoplar dos reacciones diferentes. Recuérdese que la capacidad de acoplar reacciones es una de las propiedades importantes de las enzimasInfluencia del pH sobre las velocidades de reacción enzimática
El efecto del pH sobre la velocidad de reacción de una enzima puede indicar cuáles residuos ionizables de aminoácido están en su sitio activo. La sensibilidad al pH suele reflejar una alteración en el estado de ionización de uno o más residuos que participan en la catálisis, aunque a veces se afecta la unión con el sustrato. Con mayor frecuencia, una gráfica de velocidad de reacción contra pH es una curva con forma de campana, siempre que la enzima no se desnaturalice cuando se altere el pH.

Cuando el pH es óptimo, a la mitad entre los dos valores de pKa, la mayor cantidad de moléculas de enzima está en la forma activa, con el residuo A protonado. No todas las curvas de pH-velocidad son en forma de campana. Un perfil de pH-velocidad es una curva sigmoidal si sólo participa un residuo ionizable de aminoácido en la catálisis, y puede tener una forma más complicada si participan más de dos grupos. En forma rutinaria, se analizan las enzimas cerca de su valor óptimo de pH, que se mantiene mediante el uso de soluciones amortiguadoras adecuadas.
Modos de enlazamiento en la catálisis enzimática
Son difíciles de evaluar los efectos cuantitativos de diversos mecanismos catalíticos. Ya hemos visto dos mecanismos químicos de catálisis enzimática: la catálisis ácido-base y la catálisis covalente. De acuerdo con estudios de catalizadores no enzimáticos, se estima que la catálisis ácido-base puede acelerar una reacción enzimática típica en un factor de entre 10 a 100. La catálisis covalente puede permitir la obtención del mismo aumento de velocidad.
- El efecto de proximidad
las altas concentraciones mas efectivas favorecen la formación mas frecuentes de estados de transición.
Molaridad efectiva = k1(s-1)
k2(M-1 s-1)
k1: constante de velocidad cuando los reactivos están pre-ensamblados en una sola molecular.
k2: constante de velocidad para la reacción molecular correspondiente.
- Enlazamiento débil de sustrato con enzimas

Estabilización el estado e transición
La unión de reactivos con la enzima debe ser bastante débil
- Ajuste inducido
La mayor parte de las enzimas tiene flexibilidad limitada, no son moléculas totalmente rígidas.
El ajuste inducido no es un modo catalítico, si no un efecto de especificidad del sustrato.
En 1950 Daniel Koshland sugirió que la flexibilidad de la enzima interviene en la actividad catalítica y en la especificidad del sustrato.
Su propuesta de ajuste inducido se basó en experimentos cinéticos con hexocinasa, enzima que cataliza la fosforilación de la glucosa por el ATP:
Glucosa + ATP -----> Glucosa 6-fosfato + ADP
El agua (HOH), que se parece al grupo alcohol (ROH) del C-6 en la glucosa, es lo bastante
pequeña y tiene la forma adecuada para caber en el sitio activo de la hexocinasa, por lo
que debería ser un buen sustrato.
- Estabilización del estado de transición

Algunos enzimólogos han estimado que la estabilización del estado de transición explica casi todos los aumentos de velocidad de reacción causados por las enzimas.
Gracias por la aportación en tu blog, me deja mas claro una duda que tenia con respecto a los residuos polares de los sitios activos de una enzima.
ResponderEliminar