¿Como están compuestos los carbohidratos?
- están formados por carbono, hidrógeno y oxigeno
- presentan la formula química general (CH2O).
- todos los carbohidratos presentan grupos funcionales cetona o aldehídos
- los carbohidratos se encuentran de forma abundante en las plantas y en pequeñas cantidades en animales.
- las plantas nos proveen de todos los hidratos de carbono mediante la alimentación.
Los carbohidratos pueden ser:

Aldosas: los aldehídos este grupo carbonilo se encuentra en un extremo de la cadena hidrocarbonada, por lo que tiene un átomo de hidrógeno unido a el directamente, que suele escribirse, por comodidad en la forma CHO. Según el tipo de grupo hidrocarbonado unido al grupo funcional, los aldehídos pueden ser alifáticos, R--CHO y aromáticos AR--CHO.
Cetosas: las cetonas el grupo carbonilo se encuentra unido a dos grupos hidrocarburos. Según el tipo de grupo hidrocarbonado unido al grupo funcional, las cetonas se clasifican en alifáticas, R--CO--R, aromáticas, AR--CO--AR y mixtas R--CO--AR

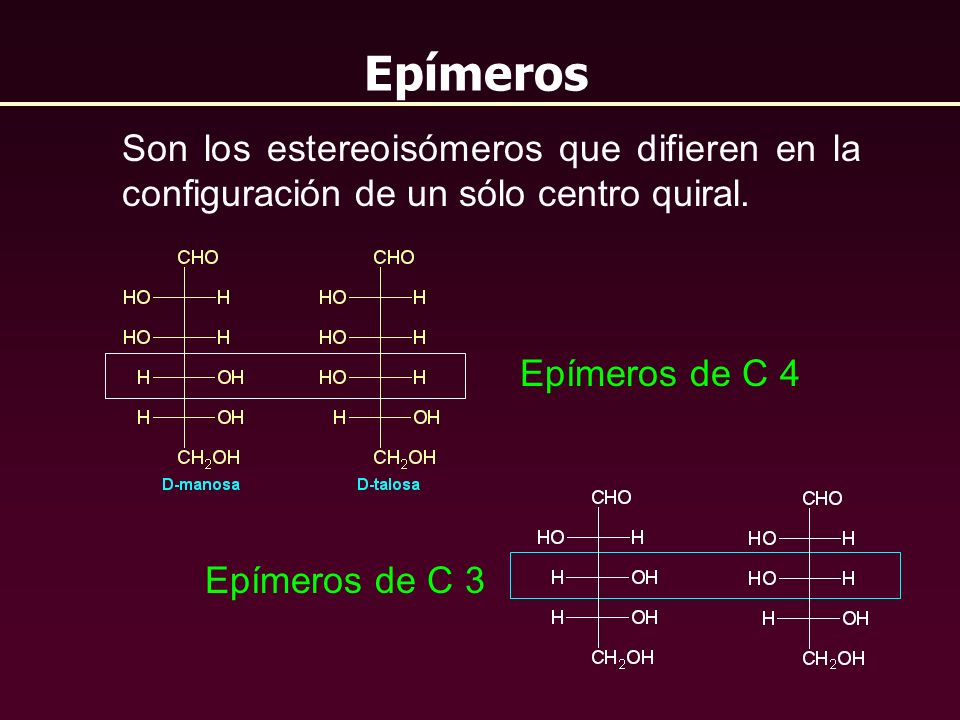
Clasificación de los carbohidratos
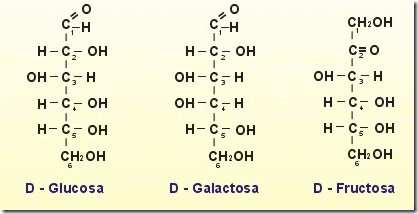
Monosacáridos
- los monosacaridos son azucares simples
- no se hidrolizan es decir que no se descomponen en otros compuestos mas simples.
- poseen de 3 a 7 atomos de carbonos
- se nombran haciendo referencia al numero de carbonos, terminando en el sufijo -osa
A. Fosfato de azucares
Los monosacáridos, en las vías metabólicas, con frecuencia se convierten en ésteres de fosfato. En la figura se muestran las estructuras de varios fosfatos de azúcar con los que se encontrará al estudiar el metabolismo de los carbohidratos. Los fosfatos de triosa, el 5-fosfato de ribosa y el 6-fosfato de glucosa son ésteres alcohol-fosfato simples. El 1-fosfato de glucosa es un fosfato de hemiacetal, más reactivo que un fosfato de alcohol. La capacidad de la UDP-glucosa para funcionar como donador de glucosilo es una prueba de esta reactividad.
B. Desoxiazucares
En la figura se muestran las estructuras de dos desoxiazúcares. En esos derivados, un átomo de hidrógeno sustituye a uno de los grupos hidroxilo del monosacárido precursor. La 2-desoxi-D-ribosa es un bloque constructivo importante en el ADN. La L-fucosa (6-desoxi-L-galactosa) está muy distribuida en plantas, animales y microorganismos. A pesar de su rara configuración L, la fucosa se deriva metabólicamente de la D-manosa.
En varios azúcares, un grupo amino sustituye uno de los grupos hidroxilo del monosacárido precursor. A veces el grupo amino está acetilado. En la figura 8.15 se ven tres ejemplos de aminoazúcares. Los aminoazúcares de la glucosa y la galactosa se suelen presentar en glucoconjugados. El ácido N-acetilneuramínico (NeuNAc) se forma a partir de la N acetilmanosamina y piruvato. Cuando este compuesto se cicla y forma una piranosa, el grupo carbonilo en C-2 (de la mitad de piruvato) reacciona con el grupo hidroxilo de C-6. El NeuNAc es un componente importante de muchas glucoproteínas y de una familia de lípidos llamados gangliósidos . El ácido neuramínico y sus derivados, incluyendo el NeuNAc, tienen el nombre colectivo de ácidos
siálicos.

D. Azucares alcoholes
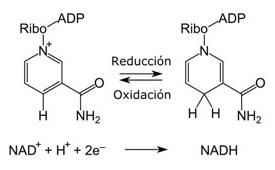
En un azúcar alcohol el oxígeno carbonílico del monosacárido precursor se ha reducido y se produce un polihidroxialcohol. La figura 8.16 muestra tres ejemplos de azúcares alcoholes. La glicerina y el mio-inositol son componentes importantes de los lípidos. El ribitol es un componente de flavina mononucleótido (FMN) y flavina adenina dinucleótido (FAD, sección 7.4). En general, los azúcares alcoholes reciben su nombre sustituyendo el sufijo -osa del monosacárido precursor por el de -itol.
E. Azucares acidos
Los azúcares ácidos son ácidos carboxílicos derivados de las aldosas, sea por oxidación de C-1 (el carbono aldehídico) para formar un ácido aldónico, o por oxidación del carbono con número mayor (el que tiene el alcohol primario) para formar un ácido aldurónico. En la figura se ven las estructuras de los derivados aldónicos y aldurónicos de la glucosa, gluconato y glucoronato. Los ácidos aldónicos existen en la forma de cadena abierta, en solución alcalina, y forman lactonas (ésteres intramoleculares) al acidularlos. Los ácidos aldónicos pueden estar como piranosas, por lo que poseen un carbono anomérico. El ácido N-acetilneuramínico (figura 8.15) es un ácido aldónico y a la vez un aminoazúcar. Los azúcares ácidos son componentes importantes de muchos polisacáridos.
El ácido L-ascórbico (figura 8.18), o vitamina C, es un enodiol de una lactona derivada del D-glucoronato. Los primates no pueden convertir glucoronato en ácido ascórbico, y en consecuencia deben obtenerlo en su dieta. El ácido ascórbico es un cofactor esencial para las enzimas que catalizan la hidroxilación de los residuos de prolina y lisina durante la síntesis de colágena
Oligosacáridos
- los oligosacaridos son uniones de 2 o mas monosacaridos.
- el enlace que une dos monosacaridos se llama enlace glicosidico.
- cuando se unen muchos monosacaridos estos son denominados polisacaridos.
 Sacarosa: La sacarosa [b-D-glucopiranosil-(1 → 2)-b-D-fructofuranósido] o azúcar de mesa es el disacárido más abundante en la naturaleza; sólo lo sintetizan las plantas. La sacarosa se distingue de los otros tres disacáridos en que su enlace glicosídico une los átomos de carbono anoméricos de dos residuos de monosacárido. Por consiguiente, las configuraciones tanto de los residuos de glucopiranosa como de fructofuranosa están fijas en la sacarosa, y ninguno de los residuos tiene libertad de equilibrarse entre sus anómeros a y b.
Sacarosa: La sacarosa [b-D-glucopiranosil-(1 → 2)-b-D-fructofuranósido] o azúcar de mesa es el disacárido más abundante en la naturaleza; sólo lo sintetizan las plantas. La sacarosa se distingue de los otros tres disacáridos en que su enlace glicosídico une los átomos de carbono anoméricos de dos residuos de monosacárido. Por consiguiente, las configuraciones tanto de los residuos de glucopiranosa como de fructofuranosa están fijas en la sacarosa, y ninguno de los residuos tiene libertad de equilibrarse entre sus anómeros a y b.
Lactosa: La lactosa [b-D-galactopiranosil-(1 → 4)-D-glucosa], carbohidrato principal en la leche, es un disacárido que sólo se sintetiza en las glándulas mamarias lactantes. Nótese que la lactosa es un epímero de la celobiosa. El anómero b de la lactosa, natural es más dulce y más soluble que el anómero b. El anómero b se puede encontrar en las nieves rancias, donde ha cristalizado durante el almacenamiento y ha dado una textura rasposa a la nieve.
 Maltosa: La maltosa es un disacárido liberado durante la hidrólisis del almidón, que es un polímero de residuos de glucosa. Existe en la malta, una mezcla que se obtiene de maíz u otros granos, que se usa en la leche malteada y en la industria cervecera. La maltosa está formada por dos residuos de glucosa unidos por un enlace a-glicosídico.
Maltosa: La maltosa es un disacárido liberado durante la hidrólisis del almidón, que es un polímero de residuos de glucosa. Existe en la malta, una mezcla que se obtiene de maíz u otros granos, que se usa en la leche malteada y en la industria cervecera. La maltosa está formada por dos residuos de glucosa unidos por un enlace a-glicosídico.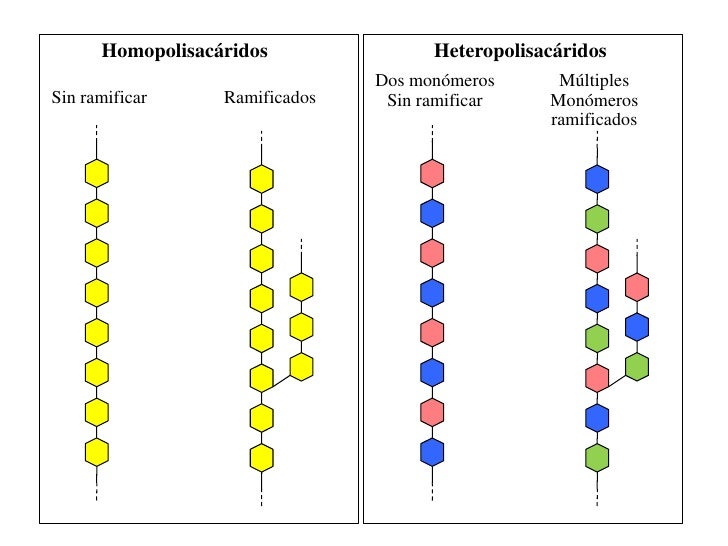 Polisacáridos
Polisacáridos
se clasifica según su composición en:
Homopolisacaridos: son polímeros que sólo contienen residuos de un tipo de monosacárido.Heteropolisacaridos: son polímeros que contienen residuos de más de un tipo de monosacárido.
se clasifica según su función en:
polisacáridos de reserva
- almidón: En las células vegetales, el almidón existe como mezcla de amilosa y amilopectina, y se almacena en granos cuyos diámetros van de 3 a 100 mm.
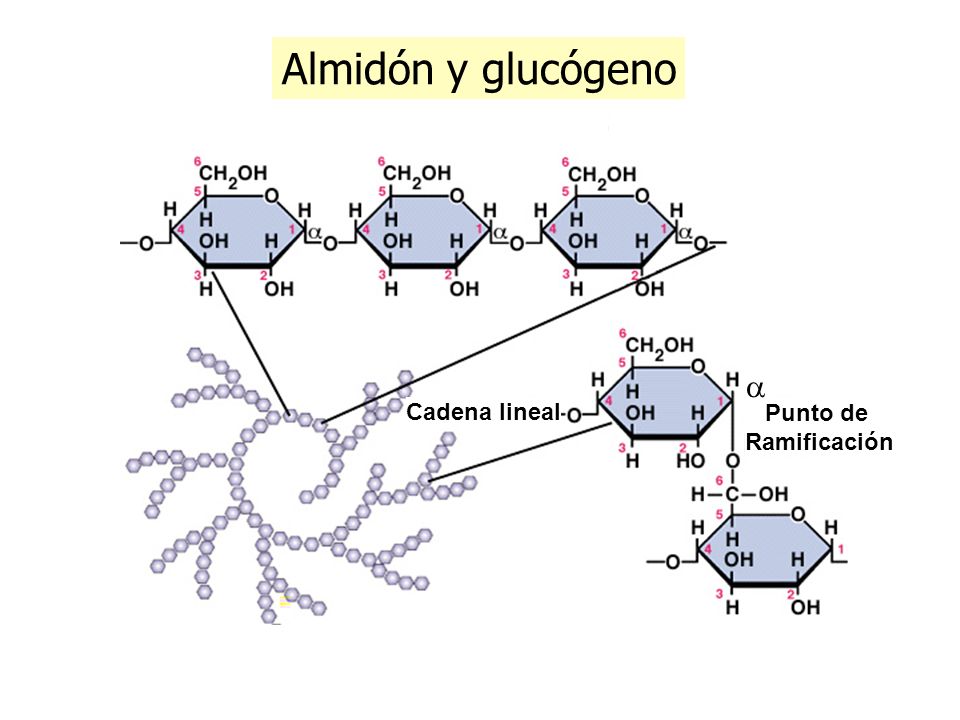
- glucógeno: el glucógeno es un polímero ramificado de residuos de glucosa. Contiene los mismos tipos de enlace de la amilopectina, pero en el glucógeno las ramas son más pequeñas y más frecuentes, y se presentan cada ocho a 12 residuos. En general, las moléculas de glucógeno son mayores que las de almidón, y contienen hasta unos 50 000 residuos de glucosa.
- celulosa: es un polisacárido estructural. Es uno de los principales componentes de las paredes celulares rígidas que rodean muchas células vegetales. Los tallos y las ramas de muchas plantas están formados principalmente por celulosa. Este solo polisacárido forma un porcentaje apreciable de toda la materia orgánica en la Tierra.
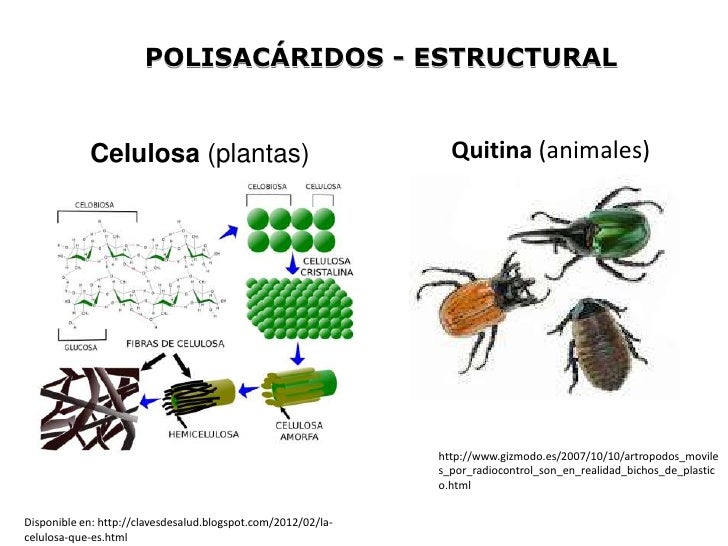
- quitina: tal vez el segundo compuesto más abundante en la Tierra, es un homoglicano estructural que se encuentra en los exoesqueletos de los insectos y crustáceos, y también en las paredes celulares de la mayor parte de los hongos y en muchas algas. La quitina es un polímero lineal parecido a la celulosa.
Glicoconjugados
Los glicoconjugados consisten en polisacáridos unidos a (conjugados con) proteínas o péptidos. En muchos casos, los polisacáridos consisten en varias unidades distintas de monosacárido. Por consiguiente, son heteroglicanos. (Almidón, glucógeno, celulosa y quitina son homoglicanos). A. Proteoglicanos: son complejos de proteínas y una clase de polisacáridos llamados glicosaminoglicanos. Esos glicoconjugados se presentan principalmente en la matriz extracelular (tejido conectivo) de animales multicelulares.
A. Proteoglicanos: son complejos de proteínas y una clase de polisacáridos llamados glicosaminoglicanos. Esos glicoconjugados se presentan principalmente en la matriz extracelular (tejido conectivo) de animales multicelulares.Los glicosaminoglicanos son heteroglicanos no ramificados de unidades repetitivas de disacárido.
 B. Peptidoglicanos: son polisacáridos unidos a péptidos pequeños. Las paredes celulares de muchas bacterias contienen una clase especial de peptidoglicano con un componente de heteroglicano unido a un péptido de cuatro o cinco residuos.
B. Peptidoglicanos: son polisacáridos unidos a péptidos pequeños. Las paredes celulares de muchas bacterias contienen una clase especial de peptidoglicano con un componente de heteroglicano unido a un péptido de cuatro o cinco residuos.El componente peptídico de los peptidoglicanos varía entre las bacterias.
En las bacterias Gram-negativas hay una capa delgada de peptidoglicano entre la membrana plasmática interna y la membrana externa.
En las bacterias Gram-positivas no hay membrana externa, y la pared celular de peptidoglicano es mucho más gruesa
 C. Glicoproteinas: son proteínas que contienen oligosacáridos unidos en forma covalente (es decir, proteínas que están glicosiladas; los proteoglicanos son un tipo de glicoproteína). Las cadenas de carbohidrato de una glicoproteína varían de longitud, de 1 hasta más de 30 residuos, y pueden formar hasta 80% de la masa total de la molécula. Las glicoproteínas son un grupo extraordinariamente diverso de proteínas que abarca enzimas, hormonas, proteínas estructurales y proteínas de transporte.
C. Glicoproteinas: son proteínas que contienen oligosacáridos unidos en forma covalente (es decir, proteínas que están glicosiladas; los proteoglicanos son un tipo de glicoproteína). Las cadenas de carbohidrato de una glicoproteína varían de longitud, de 1 hasta más de 30 residuos, y pueden formar hasta 80% de la masa total de la molécula. Las glicoproteínas son un grupo extraordinariamente diverso de proteínas que abarca enzimas, hormonas, proteínas estructurales y proteínas de transporte.
Medicamentos con proteoglicanos



Suplemento que contiene carbohidrato.

Uso
Es un ganador de peso el cual es utilizado como suplemento
para masa muscular, aportan una cantidad de calorías muy elevada, que puede ir
de 1000 a 2000 calorías por servicio, y pueden formar parte de la dieta para
ganar masa muscular. Es utilizado por deportistas que practican disciplinas
como heterofilia, fisiculturismo, por personas que desean aumentar su volumen
corporal, etc.
Dosis e
Indicaciones de Uso
Para llegar Masa tecnología Extreme 2000:
Mezclar 1 porción (6 cucharadas soperas) de 16 a 20 oz de agua o leche
descremada en una licuadora y consumen una vez al día. Alternativamente,
mezclar 1/2 taza (3 cucharadas soperas) con 8 a 10 oz de agua o leche
descremada y consumen dos veces al día. Tomar por la mañana, entre las comidas,
o después del entrenamiento. Lea toda la etiqueta antes de usar y siga las
instrucciones que se facilitan. Beba de 8 a 10 vasos de agua al día para una
buena salud general.
Nombre
del carbohidrato:
Isomaltulosa
Su nombre
químico es: 6-0-α-D-glucopyranosyl-D-fructose
Su fórmula
química es: (C12H22O11)

Isomaltulosa, también conocida comercialmente como
Byaltulosa, es un disacárido que se obtiene comercialmente por la conversión
enzimática de sacarosa a través de fermentación bacteriana
Características
La Isomaltulosa se presenta de forma natural en la caña de
azúcar y en la miel de abeja. Presenta propiedades organolépticas muy parecidas
a la sacarosa, además contiene su mismo valor calórico (4kcal/g). Se ha
utilizado como sustituto de la sacarosa en diferentes alimentos sin poder
detecar en estos ningún cambio organoléptico. La isomaltulosa no es
cariogénica, es decir no produce caries, además, no incrementa la glucosa en la
sangre ni los niveles de insulina en el consumidor. La isomaltulosa presenta
una muy baja velocidad de hidrólisis y absorción en el cuerpo comparado con la
sacarosa y otros azúcares por lo que se utiliza como fuente de energía
prolongada. La isomaltulosa se absorbe totalmente como glucosa y fructosa en el
intestino delgado. Debido a estas y otras características ha sido ampliamente
aceptado por los fabricantes de productos para diabéticos, productos libres de
azúcar, bebidas energéticas, suplementos alimenticios, farmacéuticos, dulces,
chocolates, snacks, entre otros.












{kind=link}